Una estimación estadística de la aparición de inteligencia extraterrestre en la Vía Láctea


1
Grado 12, Santiago High School, Corona, CA 92881, EE. UU.
2
Laboratorio de propulsión a chorro, Instituto de Tecnología de California, Pasadena, CA 91109, EE. UU.
3
División de Ciencias Geológicas y Planetarias, Instituto de Tecnología de California, Pasadena, CA 91125, EE. UU.
*
Autor a quien debe dirigirse la correspondencia.
Galaxias 2021 , 9 (1), 5; https://doi.org/10.3390/galaxies9010005
Recibido: 13 de diciembre de 2020 / Revisado: 11 de enero de 2021 / Aceptado: 12 de enero de 2021 / Publicado: 18 de enero de 2021
En el campo de la astrobiología, la ubicación precisa, la prevalencia y la edad de la inteligencia extraterrestre potencial (ETI) no se han explorado explícitamente. Aquí, abordamos estas preguntas utilizando un modelo de simulación galáctica empírico para analizar las variaciones espacio-temporales y la prevalencia de ETI potencial dentro de la Galaxia. Este modelo estima la ocurrencia de ETI, proporcionando orientación sobre dónde buscar vida inteligente en la Búsqueda de ETI (SETI) con un conjunto de criterios, que incluyen propiedades astrofísicas bien establecidas de la Vía Láctea. Además, se incorporan factores que normalmente se pasan por alto, como el proceso de abiogénesis, las diferentes escalas de tiempo evolutivas y la autoaniquilación potencial, para explorar la propensión al crecimiento de la ETI. Examinamos tres parámetros principales: (1) la tasa de probabilidad de abiogénesis (λ A); (2) escalas de tiempo evolutivas (T evo ); y (3) probabilidad de autoaniquilación de la vida compleja (P ann ). Descubrimos que P ann es el parámetro más influyente que determina la cantidad y la edad de la vida inteligente galáctica. Nuestra simulación de modelo también identificó una ubicación máxima para ETI en una región anular aproximadamente a 4 kpc del centro galáctico alrededor de 8 mil millones de años (Gyrs), con vida compleja disminuyendo temporal y espacialmente desde el punto máximo, afirmando una alta probabilidad de vida inteligente en el disco interior galáctico. Las distribuciones de edad simuladas también sugieren que la mayor parte de la vida inteligente en nuestra galaxia es joven, lo que dificulta la observación o detección.
Palabras llave : Fermi Paradox ; inteligencia extraterrestre ; Via Láctea
1. Introducción
Durante siglos, ha habido preguntas fundamentales sobre la vida inteligente dentro de la Galaxia. El estudio de la inteligencia de vida extraterrestre (ETI) se ha basado durante mucho tiempo en una sola muestra: la Tierra. La búsqueda de ETI (SETI, por ejemplo, [ 1 ]) ha estado muy limitada por nuestra tecnología actual y una falta de comprensión cuantitativa de la vida inteligente a escala galáctica, y las respuestas a la ausencia de descubrimiento de vida en otros lugares han sido controvertidas. Por ejemplo, un estudio anterior [ 2 ] sugirió que la paradoja de Fermi resulta de la falta de evidencia y estimaciones adecuadas de ETI. Sin embargo, muchos académicos cuestionan esta postura, ya que se basó en el argumento de que SETI es lo suficientemente extenso como para excluir a ETI en otros lugares [ 3]; esto es inválido, ya que no podemos excluir a priori la posibilidad de la existencia de otra vida inteligente en la Galaxia. Un enfoque estadístico reciente también demostró que la detección de una señal de inteligencia extraterrestre es extremadamente difícil debido a limitaciones técnicas y físicas [ 4]. Por lo tanto, se justifica desarrollar un análisis espacio-temporal sobre la propensión cuantitativa de la ETI y una evaluación a escala galáctica de diferentes parámetros clave para explorar las probables características de la vida inteligente dentro de la Vía Láctea. Además, aunque depender en gran medida de la Tierra ya que una muestra puede no parecer ideal, no nos falta otra información; Con la reciente investigación de observaciones estelares, podemos aplicar nuestro conocimiento de la Tierra a planetas similares a la Tierra para desarrollar una visión general completa de dónde y cuándo la vida inteligente puede emerger y desarrollarse en la Galaxia.
Los enfoques cuantitativos adoptados para abordar este desafío se han visto influidos en gran medida por la aplicación de la ecuación de Drake [ 5 ]; una simple expresión algebraica para investigar la cantidad de vida inteligente y su naturaleza simplista la expone a muchas re-expresiones [ 6 , 7 , 8 ]. A pesar de sus beneficios, la ecuación carece de dependencia temporal [ 9 ] y no considera la evolución de las propiedades estelares de la Vía Láctea. Además, existen incertidumbres inherentes a los parámetros [ 10 , 11 ], lo que da lugar a estimaciones muy subjetivas y de amplio alcance.
La existencia de inteligencia extraterrestre está directamente relacionada con la habitabilidad y una zona habitable galáctica (GHZ); donde se encuentran los planetas habitables y donde es más probable que se forme vida potencial. Un estudio inicial [ 12 ] exploró este concepto de cinturón de vida galáctico basado en la consideración de la co-rotación de estrellas y onda de densidad. Posteriormente, otro estudio [ 13 ] propuso aspectos espaciales y temporales de la habitabilidad galáctica y los evaluó cuantificando la abundancia de metales en la Galaxia y prediciendo la favorabilidad para la formación de planetas. Esta idea se amplió [ 14] para incluir discusiones sobre eventos catastróficos de radiación transitoria, la formación de un Júpiter caliente, y estimó que GHZ es una región anular de radio galactocéntrico entre 7-9 kpc que se ensancha con el tiempo. Además, con los mismos requisitos previos para la vida compleja que el estudio anterior [ 14 ], se desarrolló una simulación de Monte Carlo [ 15 ] de planetas habitables que muestreaba estrellas individuales para investigar dónde se encuentran las estrellas habitables, adoptando los atributos estelares galácticos de un interior. historia de formación estelar [ 16 ] y una densidad de números estelares tridimensionales [ 17]. Basándose en este modelo, predijeron que es probable que la mayoría de los planetas habitables se encuentren en el interior de la Galaxia, con una fracción mayor de estrellas albergando un planeta habitable. Sin embargo, una perspectiva alternativa sugirió [ 18 ] que todo el disco de la Galaxia puede ser habitable, ya que no se han confirmado las condiciones necesarias para que la Galaxia interior sea habitable.
Además, inspirados por la investigación anterior sobre habitabilidad, las investigaciones sobre la vida inteligente dentro de la Vía Láctea se han desarrollado con técnicas de Monte Carlo. Ampliando el modelo de Monte Carlo [ 15 ], una nueva simulación [ 19 ] exploró las oportunidades de ETI al considerar la escala de tiempo extendida requerida para que la vida evolucione hacia la inteligencia. Este estudio postuló que las brechas entre los eventos de radiación transitoria que permiten la evolución pueden variar mucho en función de una serie de factores espacio-temporales. Respaldando los resultados anteriores [ 15], este trabajo concluyó que la mayor cantidad de oportunidades ocurren en el interior de la Galaxia a pesar de una mayor frecuencia de eventos de esterilización. Sin embargo, no pudo proporcionar una estimación precisa de la ubicación del pico dentro de su rango galáctico limitado. De manera similar, otros estudios [ 8 , 20 ] describieron un modelo de Monte Carlo que estima la probabilidad de que la vida en un planeta habitable evolucione hacia la inteligencia mediante la adopción de un modelo simplificado que restablece la vida en intervalos regulares. Otro estudio [ 21 ] utilizó una distribución gaussiana para modelar la prevalencia de la inteligencia y los tiempos entre llegadas de la inteligencia sucesiva; sin embargo, también carecía de análisis espacial y temporal.
En general, los estudios previos de habitabilidad y la probabilidad de vida inteligente han proporcionado muchas ideas valiosas para la práctica de SETI; sin embargo, dadas las muchas incertidumbres involucradas, la propensión precisa de la vida inteligente galáctica a emerger aún no ha sido explorada con análisis espacial y temporal, ni ninguna investigación ha estimado explícitamente una distribución de edad para la vida potencial dentro de la Galaxia. Además, tres componentes principales han permanecido prácticamente intactos: (1) la probabilidad de vida en una condición prebiótica, (2) diferentes escalas de tiempo potenciales para la evolución biológica y (3) la probabilidad de autoaniquilación de la vida compleja. El trabajo presentado aquí incorporará estos componentes importantes, aunque a menudo descuidados, en nuestro modelo galáctico.
Para abordar nuestras preguntas sobre el surgimiento de la vida, la forma más directa y prometedora es buscar vida en otros planetas dentro y fuera del Sistema Solar. Con observaciones estelares recientes y poder computacional, es posible un enfoque teórico. Inspirado en trabajos anteriores [ 22 , 23 , 24 , 25], tratamos el proceso de abiogénesis como un proceso de Poisson para adquirir la probabilidad de que la vida surja en una condición prebiótica como un producto del tiempo y una tasa uniforme (λ). Usando nuestro modelo, si evaluamos cuánto cambia la cantidad de vida compleja en la Vía Láctea dentro de un rango de tiempo específico, podríamos tener una mejor comprensión de la hipótesis de los orígenes de la vida. Quizás podamos acercarnos a responder algunas de las preguntas más filosóficas de la humanidad, como la Hipótesis de las tierras raras.
Se ha asumido en gran medida que la escala de tiempo para la evolución biológica es idéntica a la de las especies intelectuales de la Tierra (por ejemplo, [ 14 , 15 ]). Sin considerar la posibilidad de que la vida requiera diversas escalas de tiempo para evolucionar (componente 2), cualquier resultado de la propensión al crecimiento de la vida depende del debate en curso sobre si los humanos en la Tierra son o no el paradigma que se asemeja a todas las demás formas de vida complejas en evolución. Muchos estudiosos advierten contra considerar esto como una escala de tiempo típica para la evolución (por ejemplo, [ 26 , 27 ]). Para evaluar esta incertidumbre, seleccionamos diferentes escalas de tiempo potenciales y presentamos los resultados variantes para explorar la influencia de este parámetro y todos los resultados posibles.
Además, las posibilidades potenciales de autoaniquilación durante el desarrollo de la vida compleja (componente 3) se han ignorado en gran medida (p. Ej., [ 19 ]). Aunque algunos estudios han intentado incluir esta perspectiva [ 8 , 2%0 ], no ha habido un examen galáctico sobre la influencia de este parámetro en la cantidad de ETI con consideraciones espacio-temporales; por lo tanto, la naturaleza subjetiva de este tema permanece intacta.
Las cuestiones de la ETI deben abordarse de una manera más cuantitativa y científica, teniendo en cuenta las perspectivas tanto astrofísicas como biológicas para explorar el surgimiento, el desarrollo, la prevalencia y la distribución por edades de la vida compleja en la Vía Láctea a lo largo del tiempo y la ubicación, y proporcionar una Complemento a SETI con líneas directas de investigación. Al explorar la prevalencia espacio-temporal de la vida inteligente con los parámetros presentados anteriormente, no solo permite reducir las especulaciones, sino que también mide las posibles resoluciones de la paradoja de Fermi.
2. Metodología
Desarrollamos un modelo lagrangiano con técnicas de Monte Carlo para simular con precisión cada planeta similar a la Tierra en la Vía Láctea. Este modelo incorporó un marco de especificaciones Rust ECS (Entity-Component-System) para administrar eficazmente cada partícula de estrella y planeta, y una técnica de hash espacial para optimizar la simulación a gran escala en tiempo real.
El método de simulación general se resume a continuación:
- Se inició una tabla hash espacial en 3D de la Vía Láctea con masa de gas distribuida;
- Estrellas similares al Sol generadas que albergan planetas similares a la Tierra y explosión de supernova activada con la misma distribución que las observaciones;
- Para cada planeta similar a la Tierra, permita que la vida emerja con el proceso de abiogénesis de Poisson;
- Para cada planeta portador de vida libre de eventos transitorios (por ejemplo, supernova), siga la evolución de la vida hacia la inteligencia.
2.1. Formación de una estrella similar al Sol que alberga un planeta similar a la Tierra
2.1.1. Prevalencia de estrellas similares al Sol que albergan planetas similares a la Tierra
Dentro de nuestra simulación, definimos estrellas similares al Sol como estrellas con masa de 0.8 M ⊙ ≤ M ≤ 1.2 M ⊙ , donde M ⊙ = 1 denota la masa del Sol. También adoptamos la función de masa inicial (IMF, la distribución inicial de masa para una población de estrellas), con α = 2,35 [ 28 ], y calculamos la fracción de estrella similar al Sol aproximada en la Vía Láctea. A continuación, describimos los planetas similares a la Tierra como planetas: (1) con 1-2 radios terrestres, (2) recibiendo energía estelar dentro de un factor de 4 en comparación con el de la Tierra y (3) con un período orbital similar a la Tierra de 200 a 400 días. Desde estas perspectivas, utilizamos análisis observacionales recientes sobre la prevalencia de planetas del tamaño de la Tierra [ 29], que estimó que el 11% de los planetas del tamaño de la Tierra alrededor de estrellas similares al Sol reciben energía estelar similar a la Tierra, y el 5.7% de los planetas del tamaño de la Tierra obtienen períodos orbitales similares a la Tierra. Multiplicamos estas probabilidades para adquirir la fracción de estrellas similares al Sol que albergan planetas similares a la Tierra en la Galaxia que se ajustan a nuestros cuatro criterios descritos anteriormente.
2.1.2. Tabla hash espacial y distribución de gas
Diseñamos una tabla hash espacial galáctica con el mismo volumen de la Vía Láctea, dividida uniformemente en celdas tridimensionales con el volumen de cada celda en 0.01 kpc 3 . Esta tabla galáctica permite el cálculo de objetos a gran escala a una velocidad de fotogramas en tiempo real y permite consultas rápidas de detección de ubicación y proximidad [ 30 ]. Un trabajo reciente [ 31 ] utilizó un enfoque similar para implementar la densidad jerárquica en la investigación de la formación y evolución galácticas.
Para simular la formación de estrellas (el colapso del gas frío bajo su propio peso en el espacio interestelar), calculamos la masa de gas para cada celda y la distribución inicial del gas en la formación de la Galaxia (1). La densidad de la superficie del gas disminuye exponencialmente con el radio galactocéntrico [ 16 ]:donde la cantidad es la densidad superficial del gas (M ⊙ kpc −2 ) y r es la distancia radial desde el Centro Galáctico (kpc). El constantees la densidad de la superficie central de la Galaxia [ 32 ], y= 2,25 kpc denota la longitud de escala del disco radial. En el momento de la formación de la galaxia simulada, esta ecuación asigna diferentes densidades superficiales totales de gas a cada celda en función de su distancia del Centro Galáctico.
2.1.3. Límite superior de formación estelar
Para permitir la formación de estrellas a partir del gas interestelar, utilizamos la ley de Schmidt-Kennicutt [ 33 ], que es una relación simple entre la densidad de la superficie del gas y la densidad de la superficie de la tasa de formación de estrellas (SFR):donde la cantidad de ∑ SFR representa la densidad superficial de SFR (M ⊙ kpc −2 Myr −1 ), y es lo adimensional de la Ecuación (1) en la unidad de M ⊙ kpc −2 . La constante A = 250 M ⊙ kpc −2 Myr −1 [ 34 ] y N = 1.4 [ 34 , 35 ]. Para cada celda dentro del modelo galáctico, primero calculamos su desde el asignado inicialmente descrito anteriormente, y luego calculó la masa estelar teórica de SFR (M SFR , en M ⊙ Myr −1 ) multiplicando su área de superficie. Estimamos la M SFR para estrellas similares al Sol que albergan planetas similares a la Tierra de acuerdo con la fracción de la Sección 2.2 . Para cada paso de tiempo (Myr), nuestro modelo galáctico acumula la masa estelar teórica disponible para la formación de estrellas (M SF , en M ⊙ ). Esto significa que, en cada ubicación, la masa estelar recién disponible del colapso del gas se agregará al M SF total disponible de acuerdo con el M SFR.. Luego adquirimos la masa estelar teórica para la formación de estrellas y la usamos como límite superior para la masa estelar total de formación de estrellas en cada celda dentro del modelo galáctico. Esta estimación teórica no está destinada a la SFR, sino más bien, a incorporar la relación entre la formación de gas y estrellas. Utilizamos el SFR de acuerdo con un modelo de evolución química reciente de la historia de formación de estrellas (SFH), y el proceso de formación de estrellas se describirá en detalle en la Sección 2.1.5 .
2.1.4. Masa estelar y vida útil de la secuencia principal
Para cada estrella, asignamos uniformemente los rangos de masa de 0,8 M ⊙ ≤ M ≤ 1,2 M ⊙ y su vida de secuencia principal, la vida útil total de la estrella, correspondiente a su masa [ 36 ]:dónde = 11.000 es la vida de la secuencia principal del Sol en Myr [ 37 ]. Cuando una estrella envejece hasta su secuencia principal de vida, la masa estelar de la estrella se devuelve al M SF total disponible en la celda correspondiente, de forma análoga a la formación de nebulosas planetarias.
2.1.5. Modelo de formación estelar
Utilizamos un modelo de evolución química desarrollado previamente de SFH [ 38 ], que derivó SFH de abundancias estelares ajustando las tendencias de abundancia con la edad en lugar del procedimiento habitual de fijar la prescripción de formación de estrellas. Esto permite una precisión sin precedentes de la historia de la formación de estrellas de la Galaxia en los primeros mil millones de años, según se muestreó en la vecindad solar. Su modelo obtuvo dos fases distintas de la formación del disco interior galáctico (con un radio galactocéntrico <8 kpc): la formación de disco grueso y delgado, y la formación del disco exterior con una historia igualmente plana, lo que postula que la mitad de la masa estelar de la estrella se forma en el disco grueso de la Vía Láctea aproximadamente a 4 Gyrs después de su formación.
Preferimos este enfoque porque este modelo incorporó explícitamente la importancia del disco galáctico grueso, un componente que fue reconocido ya en 1986 [ 39 ]. Mientras que el disco grueso representa la población estelar galáctica en la fase más intensa de formación estelar [ 40 , 41 ], se ignora en gran parte en muchos modelos de formación estelar cuando se usa la vecindad solar como una restricción que limitaba las metalicidades intermedias de las estrellas. Por ejemplo, los modelos anteriores [ 16 , 42 ] carecían de la consideración de la fase explícita de disco grueso, lo que resultaba en la metalicidad de la masa estelar estimada un 10% menos que la masa estelar total.
Usando la técnica de Realización de Monte Carlo, permitimos que cada estrella se formara estocásticamente en cada paso de tiempo en las respectivas regiones del disco (interior, exterior), y calculamos el SFR correspondiente del modelo SFH [ 38 ]. En cada paso de tiempo (Myr), permitimos que las estrellas se generen en un bucle de acuerdo con el SFR correspondiente, con cada estrella asignada a una coordenada aleatoria de ( X , Y , Z ). Luego, nuestro modelo obtuvo el M SF calculado (descrito en la Sección 2.1.3 ) de la celda correspondiente en esa ubicación, y verifica si hay suficiente masa estelar disponible para que se forme la estrella. Si se cumplen los criterios, el total de M SFen esa celda se restará la masa estelar de la estrella recién formada, asemejándose a la conversión de gas en estrellas. Este proceso de formación de estrellas se repetirá a través del bucle del SFR del modelo SFH, variando según las diferentes fases temporales de la Galaxia. Al combinar el enfoque teórico de la prescripción de formación estelar de la ley de Schmidt-Kennicutt con un modelo SFH derivado de las observaciones estelares precisas, nuestro modelo tiene mayor poder, ya que respeta la naturaleza de la formación estelar y no depende demasiado de un solo modelo. .
2.2. Supernova
La tasa de explosión de supernova (SNe) es uno de los principales componentes de los modelos habitables [ 14 , 15 ]. Un entorno habitable se describe a menudo como una condición que no se expone con frecuencia a eventos transitorios, como SNe. Simulamos SNe, utilizando la frecuencia de SN galáctica estimada por un estudio previo [ 43 ], en el que se modeló una tasa de SNe de 2,5 × 10 -2 años -1 y se concluyó que el 85% de estos SNes son de progenitores masivos. Esto es consistente con las estimaciones teóricas [ 44 ] que predijeron que la velocidad de la supernova Tipo II (SNII, explosión de una estrella con una masa superior a 8 M ⊙ ) sería de 1,96-3,35 × 10 -2 años -1. La frecuencia de eventos de supernova de Tipo Ia (SNIa, explosión de una enana blanca en un sistema binario) es estimada por un estudio anterior [ 45 ], que encontró el valor de 2.25 a 2.9 × 10 −3 años −1 .
Se investigó el efecto de extinción de un SNII promediado [ 46 ], que predijo que a una distancia dentro de 0.008 kpc, el flujo de un evento SNII es suficiente para agotar la capa de ozono de cualquier planeta cercano y así esterilizar cualquier vida terrestre en ese planeta. A partir de esta estimación, asignamos la distancia de esterilización para cada SNII y SNIa de acuerdo con su distribución de magnitud absoluta. Con 46 muestras SNII observadas de una galaxia externa, un estudio anterior [ 47 ] abordó la distribución de SNII y concluyó que una magnitud SNII promedio en la banda de longitud de onda B (M B , aproximadamente 435 nm de longitud de onda) era de -16,89 y σ = 1,35. Además, un estudio reciente [ 48] desarrolló la distribución SNIa a partir de 239 muestras SNIa con correcciones K y un proceso de corrección de sesgo, que encontró la media M B de −19,25 y σ = 0,5. A partir de estas distribuciones de SN, proponemos una parametrización simple para la distancia de esterilización de cada SNII y SNIa, que se inspira en el enfoque del trabajo anterior [ 15 ]:donde D SNe es la distancia que resultará en la extinción de la vida de un SN dado a un planeta portador de vida cercano, y M SN es la magnitud absoluta del SN dado. La constante d SNII = 0,008 kpc denota la distancia de extinción de un SNII medio [ 46 ], y M SNII = –16,89 es la magnitud absoluta media M B de SNII [ 47 ]. Reconocemos que esta parametrización puede no ser la más realista, considerando la complejidad de los mecanismos de esterilización causados por SNe; sin embargo, es bastante razonable modelar SNe en nuestra simulación.
Cuando ocurre un evento de esterilización en un planeta portador de vida, la capa de ozono de ese planeta se agota y la vida terrestre dentro del planeta se elimina. En nuestra simulación, permitimos que la vida se repitiera dado un cierto período de tiempo como T min; el surgimiento de la vida se define como un proceso de abiogénesis de Poisson, que se presenta en la siguiente subsección.
2.3. Proceso de Poisson de abiogénesis
La abiogénesis, la creación de vida a partir de sustancias inanimadas, se considera el camino más probable hacia el surgimiento de la vida [ 49 ]. Usando modelos matemáticos, asumimos que la vida es posible para estrellas similares al Sol que albergan planetas similares a la Tierra, en los que la vida emerge con el proceso de abiogénesis de Poisson. En este proceso, asumimos que los eventos separados de abiogénesis no se influyen entre sí, y que la probabilidad de abiogénesis es simplemente un producto de una tasa uniforme (λ A , con unidad Myr −1 ) y el tiempo. Además, interpretamos la química compleja y de múltiples caminos hacia la acumulación de vida como un conjunto, similar al trabajo anterior (por ejemplo, [ 22 ]). Por tanto, la probabilidad de que surja vida en un planeta dentro del tiempo T es:donde P Poisson [λ A , T, n = 0] es la probabilidad de obtener cero eventos exitosos, λ A es el parámetro de tasa, o la tasa de probabilidad de abiogénesis en un planeta similar a la Tierra por unidad de tiempo (Myr), y T min es el período de tiempo en el que un planeta joven parecido a la Tierra tenía un entorno severo que impedía la vida en el momento de su formación, elegido uniformemente en un rango de 0,1 a 1 Gyr. Preferimos este enfoque de T minsobre una distribución normal o un valor elegido específico porque proporcionará un valor relativamente imparcial y permitirá que varíen diferentes planetas similares a la Tierra, ya que el período de tiempo que excluye estrictamente la vida en la Tierra sigue siendo controvertido. Reconocemos que este rango en sí mismo es una suposición; sin embargo, como actualmente carecemos de una mejor comprensión del origen de la vida, este rango es lo suficientemente razonable como para limitar los resultados y permitir que ocurran algunas variaciones.
Con esta simplicidad, nos damos cuenta de una debilidad importante: el parámetro λ A puede no ser una constante uniforme incluso cuando los planetas son todos similares a la Tierra y posiblemente comparten mecanismos comunes para el desarrollo de la vida. Las disparidades de los entornos planetarios a lo largo del tiempo también pueden influir en nuestros resultados. Para abordar este problema, incluimos el parámetro T min , que responde a uno de los mayores cambios de un ambiente severo a uno más leve. Argumentamos que, dado que la mayoría de los cambios marcados en la Tierra provienen de las actividades de la vida después del primer éxito de un evento de abiogénesis, esos cambios son irrelevantes para este proceso de nuestro modelo. Además, aunque un estudio reciente [ 23 ] sugirió que la tasa de abiogénesis, λ A, Depende en gran medida de las condiciones ambientales específicas, suponemos que la Tierra-como planetas probable que tenga un ritmo muy similar de la abiogénesis, λ son Una .
Aunque el trabajo anterior intenta proporcionar algunas estimaciones de λ A (por ejemplo, [ 22 ]), notamos que el valor de este parámetro varía mucho. Para ampliar el trabajo previo de cuantificación de λ A , no asumimos ningún valor en particular, sino que simplemente presentamos resultados variables con diferentes valores de λ A y evaluamos sus pesos en una escala global de sistemas estelares distribuidos. Elegimos que el rango de valores del parámetro fuera 1 y 10 −6 Myr −1 , los límites superior e inferior obtenidos del rango sugerido por el estudio anterior [ 22 ].
2.4. Tiempo suficiente para la evolución de la inteligencia
No tenemos pruebas suficientes para atribuir un valor específico a la escala de tiempo necesaria para que la vida se convierta en inteligencia (denotado como T evo , en Gyr). Si bien la Tierra puede proporcionar una inferencia hipotética para este parámetro, presentamos los resultados de valores potenciales seleccionados. En la Tierra, T evo es aproximadamente de 3 a 3,5 Gyrs desde el surgimiento de la vida hasta el estado actual donde comenzamos el SETI. Se desconoce si esta es una escala de tiempo típica en otros planetas similares a la Tierra, por lo que seleccionamos tres valores potenciales: T evo ± 0.2 Gyr, donde T evo = 1, 3, 5.
Reconocemos la posible suposición hecha por los estudios anteriores [ 8 , 27 ] de que el proceso de evolución de la inteligencia consiste en subprocesos, cada uno de los cuales tiene una escala de tiempo específica. Sin embargo, a menos que se sugiera un mecanismo evolutivo particular con subprocesos con evidencia suficiente, no es necesario agregar complejidad solo para incorporar la distribución especulativa. Por lo tanto, preferimos "integrar" los posibles pasos detallados y tratar los subprocesos como un conjunto de períodos de tiempo. Además, tenemos en cuenta la explosión de SNe y sugerimos el parámetro T evoque describimos en esta subsección corresponde al período de tiempo que está libre de eventos transitorios; cuando un SNe casi esterilizante amenaza la vida terrestre durante el proceso evolutivo, simplemente reiniciamos el proceso de abiogénesis y evolución en ese planeta, permitiéndoles que se repitan a medida que el planeta envejece.
2.5. Aniquilación de la inteligencia
Si bien ninguna evidencia sugiere explícitamente que la vida inteligente eventualmente se aniquilará a sí misma, no podemos excluir a priori la posibilidad de la autoaniquilación. Ya en 1961, un estudio sugirió [ 50 ] que el progreso de la ciencia y la tecnología conducirá inevitablemente a la destrucción completa y la degeneración biológica. Esto está respaldado por muchos estudios previos que argumentan que la autoaniquilación de humanos es muy posible a través de varios escenarios (por ejemplo, [ 51 ]) que incluyen, entre otros, la guerra, el cambio climático [ 52 ] y el desarrollo de la biotecnología [ 53 ]. Además, los estudios también discutieron el impacto de fuertes restricciones de sostenibilidad en la vida de la civilización [ 54 , 55].
Para abordar estas posibilidades, desarrollamos un parámetro de probabilidad simple P ann para denotar la probabilidad de autoaniquilación de la vida del complejo galáctico. En cada paso de tiempo (1 Myr), incorporamos P ann al desarrollo de la vida compleja con métodos de Monte Carlo. Asumimos que este parámetro permanece uniformemente constante en el tiempo, lo que carece de variaciones temporales o aspectos sociales diferentes de las civilizaciones y puede resultar en una gran variación. Sin embargo, de ninguna manera intentamos estimar P ann , ya que este parámetro solo sirve para proporcionar una comprensión cualitativa de los posibles resultados de la vida inteligente, que nosotros, como vida compleja, podemos potencialmente aniquilarnos a nosotros mismos. También utilizamos diferentes valores de P annpresentar los resultados variables de escenarios de casos extremos (por ejemplo, cuando P ann se establece en 0 o 0,99) como un medio para examinar su impacto en la prevalencia global de ETI en la Vía Láctea. Probamos un rango de valores de 0 a 0,99 y seleccionamos lo siguiente para este parámetro: 0, 0,5 y 0,99; cuanto menor sea el valor de P ann, mayor será el número máximo de vida galáctica potencial. Para los tres valores, proporcionamos sus variaciones espaciales y temporales sobre la prevalencia galáctica de la vida inteligente. Excluimos el valor de 1 para el límite superior, simplemente por el hecho de que todavía estamos vivos; un valor de 1 significaría que el nivel de vida inteligente en toda la Galaxia sería cero. Por otro lado, no se puede descartar un valor de cero, ya que una civilización puede volverse inmortal [56 , 57 ].
3. Resultados y discusión
Este trabajo presenta un modelo de la Vía Láctea que simula la evolución de la ETI galáctica y ha producido un conjunto de criterios para rastrear dónde y cuándo ocurriría la vida compleja.
3.1. Análisis espacial-temporal sobre la aparición de IET
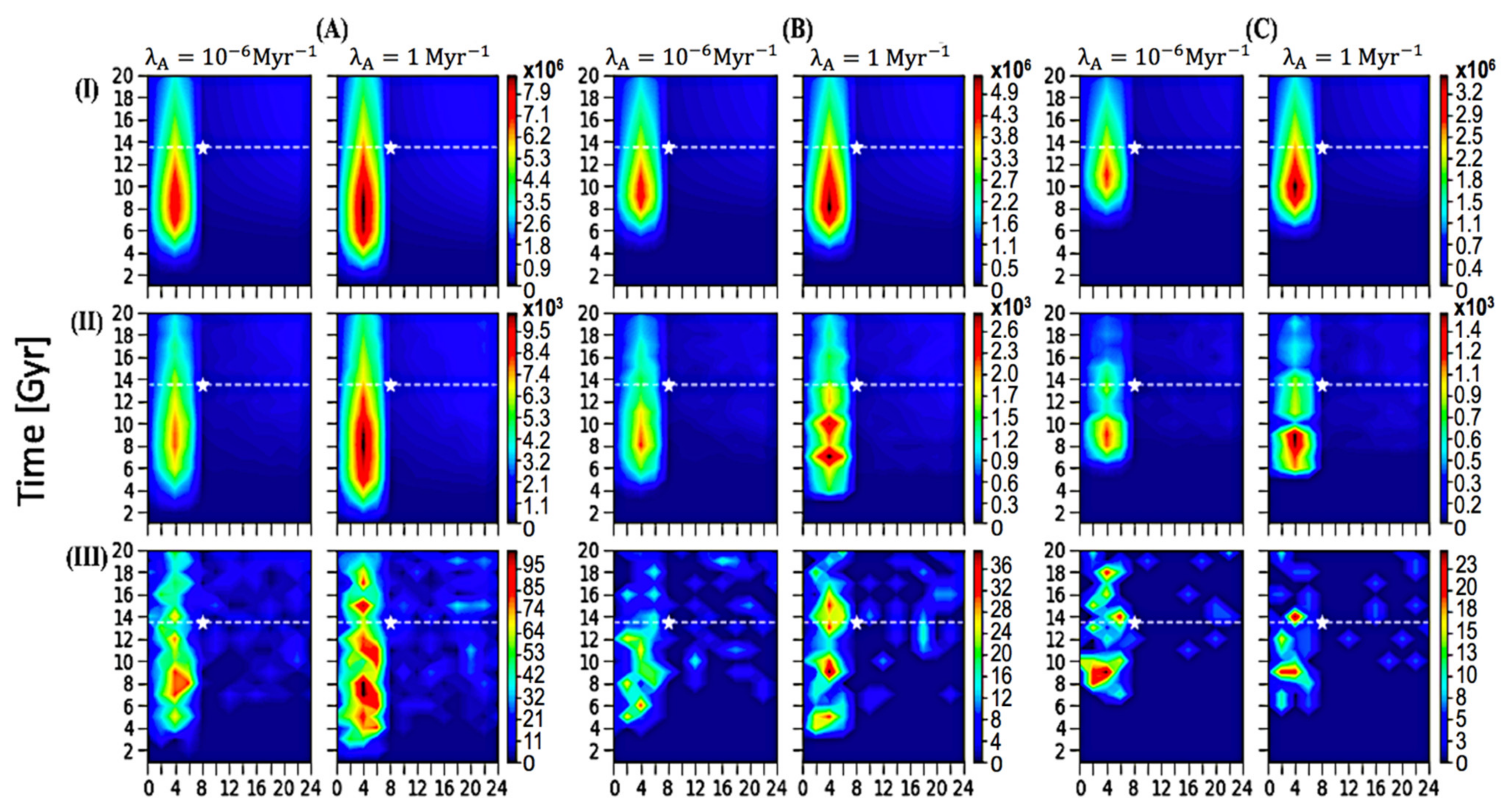
Proporcionamos 18 escenarios con resolución a escala galáctica para examinar los resultados potenciales que dependen de diversos parámetros, representados en la Figura 1 y resumidos en la Tabla 1 . Para cada parámetro clave, investigamos la cantidad de ETI (Z ETI ) con perfiles espaciales y temporales. La Figura 1 tiene la línea de tiempo actual (13 Gyrs) y nuestra ubicación en el momento actual (8 kpc, 13 Gyrs) indicadas como referencia.
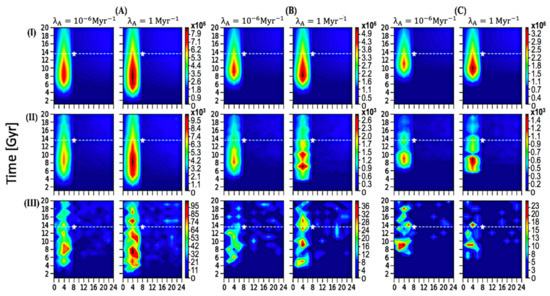
Figura 1. Perfiles espacio-temporales para Z ETI ( A ) T evo = 1 Gyr, ( B ) T evo = 3 Gyrs, ( C ) T evo = 5 Gyrs; ( I ) P ann = 0, ( II ) P ann = 0.5, ( III ) P ann = 0.99, sobre 20 Gyrs. Cada panel individual vertical corresponde a un valor específico de λ A , como se muestra en el título anterior. La estrella blanca marca la ubicación actual de nuestro sol a 8 kpc y 13,5 Gyrs, y la línea punteada blanca representa nuestra línea de tiempo actual de 13,5 Gyrs.
Tabla 1. Cada valor numérico representa la Z ETI máxima de su perfil espacio-temporal en la Figura 1 .

En la Figura 1 , se asignó un valor específico de λ A , 1 o 10 −6 Myr −1 , a cada panel vertical. Para una mejor comparación, presentamos los resultados de diferentes λ A con la misma codificación de contorno de color y encontramos resultados similares. Esto posiblemente se deba a la gran cantidad de planetas similares a la Tierra alrededor de las áreas de picos, y si se le da suficiente tiempo, la vida puede ser común allí; una muestra de vida tan significativamente grande resultó en poca variación. Por lo tanto, se sugiere que la vida está en otra parte común y la prevalencia de la ETI no depende en gran medida del parámetro λ A . Dentro del rango de tiempo desarrollado [ 22], la probabilidad de que surja la vida probablemente no sea el factor detrás de la paradoja de Fermi. A menos que asumamos especulativamente un valor para λ A muy por debajo del límite inferior del rango desarrollado por trabajos anteriores, es poco probable que el número y la prevalencia de ETI se alteren significativamente, lo que puede desacreditar la hipótesis de las tierras raras como una resolución a la paradoja de Fermi.
Como se discutió anteriormente en nuestros métodos, usamos un rango de valores de P an de 0 a 0,99 para examinar el impacto del potencial de autoaniquilación en la prevalencia de ETI. Los valores seleccionados de 0, 0,5 y 0,99 se representan en paneles horizontales (I), (II) y (III), respectivamente. Como se muestra, el rango de Z ETI varía significativamente con diferentes valores de P ann, lo que sugiere que la cantidad de vida inteligente galáctica depende en gran medida de la probabilidad de autoaniquilación. Se puede interpretar la importancia de este parámetro en la hipótesis relacionada (teoría del Gran Filtro) de que la probabilidad de autoaniquilación es posiblemente alta, lo que da como resultado una fracción extremadamente pequeña de ETI. No tomamos posición en este argumento; más bien, nuestro enfoque es resaltar la propensión al crecimiento de la vida inteligente potencial en la Vía Láctea. Observamos que en el panel (III), P ann= 0,99 muestra que algunas proporciones de las variaciones espacio-temporales se desvían de los patrones generales debido a su pequeño tamaño resultante de una posibilidad extremadamente alta de autoaniquilación; sin embargo, esto no invalidará nuestras conclusiones ya que: (1) la probabilidad de 0,99 es un escenario de caso extremo y es poco probable que la probabilidad real de autoaniquilación sea tan cercana a 1; y (2) la ubicación general del pico y cómo la prevalencia del cambio de ETI espacial y temporalmente permanece consistente en sus perfiles espacio-temporales.
Además, consideramos los resultados variables con tres T evo potenciales . Los paneles verticales de la Figura 1 A – C corresponden a una escala de tiempo de 1, 3 y 5 (0.1) Gyrs, respectivamente, cada uno representado con la misma escala de tiempo y radio galactocéntrico. En el panel de la Figura 1 A, los picos se extienden por un período de tiempo más amplio que otros dos paneles porque las civilizaciones aparecieron antes con una escala de tiempo más corta requerida para la evolución. Los resultados para la propensión al crecimiento permanecen generalmente consistentes y el rango de números no varía significativamente en comparación con lo que se presenta con diferentes valores para P ann .
Aunque el rango de Z ETI varía significativamente con el cambio en parámetros como P ann , la ubicación del pico y la propensión espacio-temporal para la prevalencia de ETI sigue siendo precisa en todos los escenarios. A partir de la Figura 1 , podemos desarrollar una imagen completa de dónde y cuándo se ha formado la vida compleja potencial: en el interior de la Galaxia entre 2 y 8 kpc, con el foco central del pico a 4 kpc del Centro Galáctico y el tiempo alrededor de 8 Gyrs, con el número disminuye monótonamente hacia afuera. El Z ETIcayó inmediatamente en el límite del disco interior galáctico (8 kpc), y permaneció con una densidad numérica baja en todo el disco exterior galáctico. Curiosamente, nuestra ubicación no está dentro de la región donde ocurre la mayoría de las ETI, ya que nuestro sol está ubicado fuera del límite de la galaxia interior (ver la estrella blanca en la Figura 1 ); nuestra ubicación puede estar simplemente demasiado lejos de otras formas de vida potencialmente complejas. Sugerimos que el SETI se dirija más hacia el interior de la Galaxia, idealmente hacia el anillo a 4 kpc del centro galáctico.
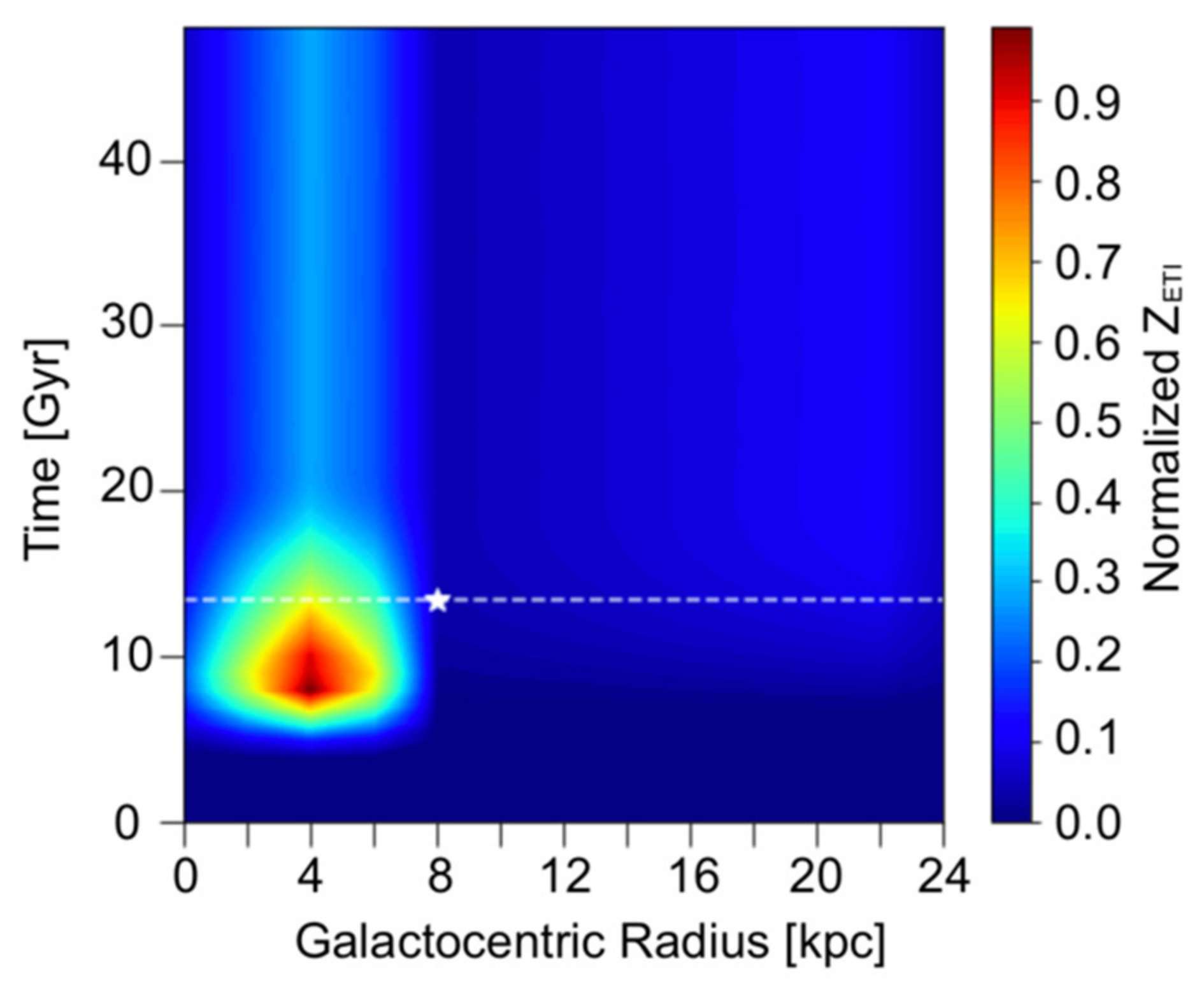
En la Figura 1 , la cantidad de vida inteligente también disminuye con el tiempo después del pico, posiblemente debido a un estallido de ETI en el momento pico, disminuyendo gradualmente a medida que las estrellas similares al Sol envejecen hasta su secuencia principal de vida, y eventualmente puede alcanzar un equilibrio entre nacimiento y muerte de la vida inteligente. En nuestro modelo, esta explicación se prueba para ser cierta en la Figura 2 trazando el cambio espacio-temporal de Z ETI normalizado en un tramo de 50 Gyrs. El parámetro P ann se establece en 0 en este gráfico, lo que sugiere que los únicos factores que dieron como resultado este equilibrio son la formación y muerte de estrellas y la esterilización de SNe. Es decir, este último estado de equilibrio también señala el punto máximo como un estallido inusual de ETI a lo largo del tiempo. En particular,La Figura 2 marca el estado de equilibrio en alrededor de 20 Gyrs, y la prevalencia de ETI se mantuvo idéntica durante los siguientes 30 Gyrs.
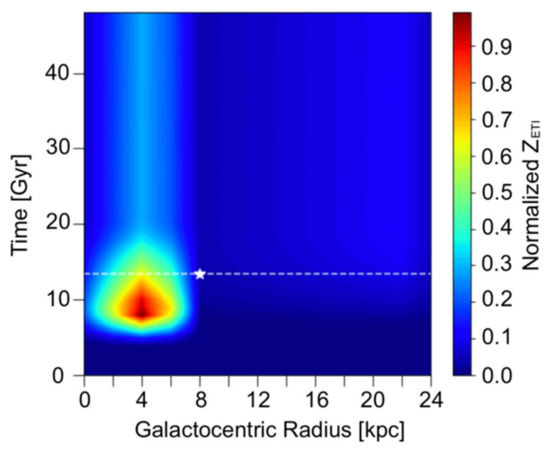
Figura 2. Perfil espacio-temporal normalizado para Z ETI sobre 50 Gyrs, cuando λ A = 10 −6 Gyr −1 , P ann = 0 y T evo = 5 Gyrs.
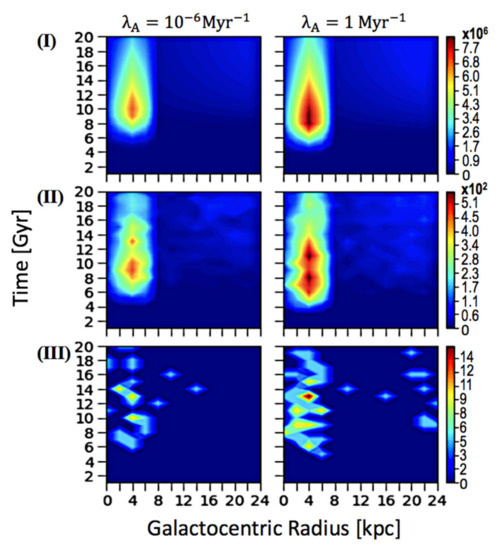
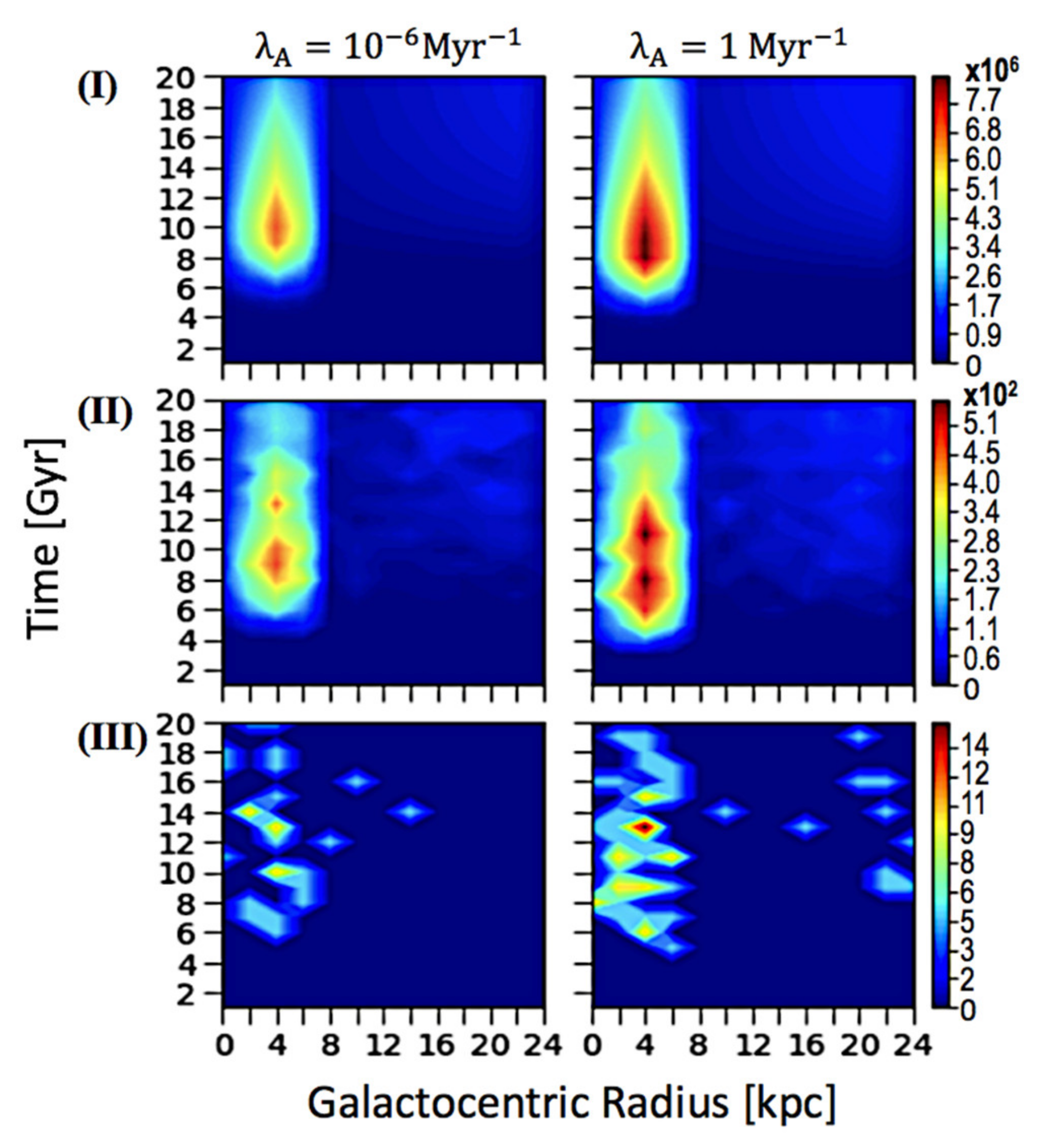
Además, para evaluar el impacto de esterilizar SNe en Z ETI , presentamos el número esterilizado de ETI con un análisis espacio-temporal similar, representado en la Figura 3 . Presentamos los resultados cuando T evo se establece en 3 Gyrs, simplemente porque una colección de parcelas es suficiente para ver la correlación entre la frecuencia de esterilización SNe y la prevalencia de ETI. Sin embargo, presentamos resultados de otros parámetros variables como un medio para comparar las parcelas con los correspondientes perfiles espacio-temporales del panel vertical Figura 1 B. Encontramos que la cantidad de ETI aumenta con la frecuencia de esterilización de SNe. Cuando el Z ETI alcanza el máximo, el evento SNe en el que la vida del complejo esterilizado también alcanza un máximo, y podemos concluir que la densidad numérica significativamente mayor de estrellas habitables supera el impacto de eventos SNe esterilizantes más frecuentes.
3.2. El efecto del parámetro de aniquilación de la inteligencia en las distribuciones de edad
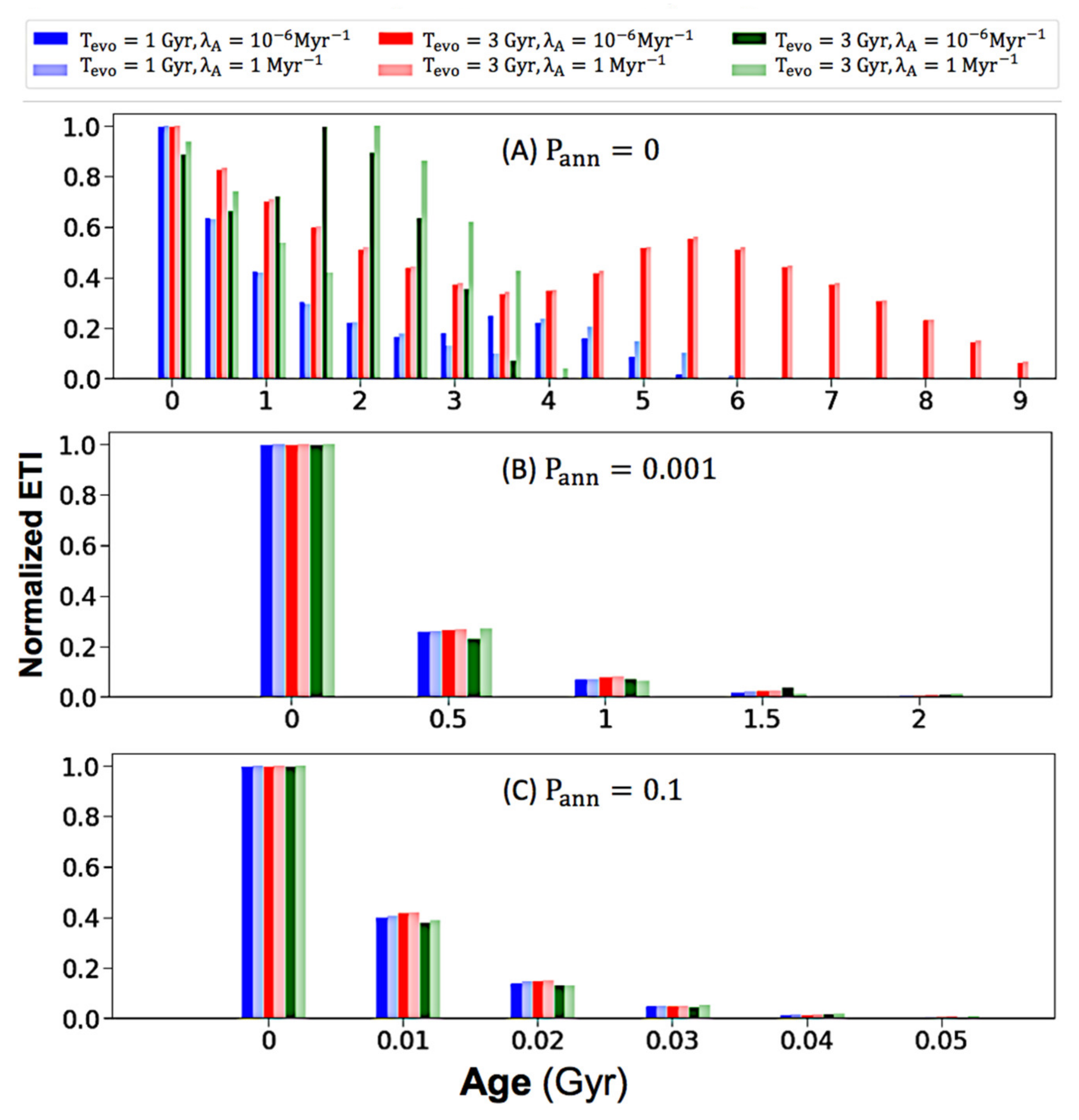
Para examinar la edad de la ETI en el momento actual de la Vía Láctea, graficamos distribuciones de edad para valores variables de los parámetros clave (T evo , λ A , P ann ), y encontramos que P ann influye de manera observable en los resultados de la distribución de edades. . Por lo tanto, ejecutamos el modelo con valores de P ann que iban de 0 a 0,99 y seleccionamos tres parcelas resultantes en la Figura 4 . Seleccionamos el valor 0, 0,001 y 0,1 porque cada uno cambia enormemente la distribución de edad. Cuando P ann se establece en 0 en el panel (a), las edades de ETI varían significativamente de 0 a 10 Gyrs. Sin embargo, esta variación cae inmediatamente cuando P annse establece en 0,001 en el panel (b), con edades que solo varían de 0 a 2,5 Gyrs. Esto sugiere que P ann debe ser extremadamente bajo para permitir que ocurran variaciones de edad. Más aún, al aumentar P ann a 0.1 en el panel de la Figura 4 C, toda la vida compleja se vuelve menor de 0.06 Gyr de edad, y la mayoría permanece menor de 0.01 Gyr. Dado que no podemos descartar la alta posibilidad de aniquilación, la Figura 4 sugiere que la mayor parte de la vida compleja potencial dentro de la Galaxia puede ser todavía muy joven.
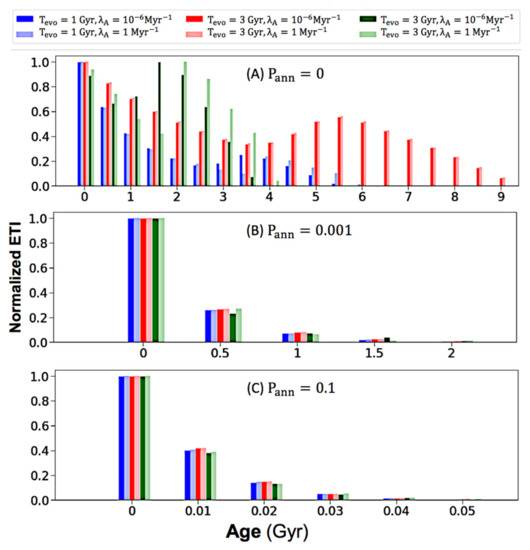
Figura 4. Distribuciones de edad ( A ) P ann = 0, ( B ) P ann = 0.001, ( C ) P ann = 0.1. El rango de cada contenedor para los paneles ( A ) y ( B ) es de 0,5 Gyr, y el rango de contenedor para el panel ( C ) es de 0,01 Gyr. Los números en las marcas del eje x representan el valor inicial de su rango de intervalo. Todos los paneles comparten la misma leyenda de codificación de colores en la parte superior.
3.3. Discusión
La ubicación exacta del pico del disco galáctico interno que sugerimos es consistente con muchos estudios previos [ 15 , 19 , 20 , 58 ]. Nuestro trabajo produjo una ubicación de pico probabilística con respecto al radio y tiempo galactocéntrico, con una resolución a escala galáctica que se extiende por más de 20 Gyrs. Este tramo de tiempo y espacio sin precedentes nos permite compilar una imagen completa de cuándo y dónde evoluciona la ETI potencial, y presenta todos los perfiles potenciales de la vida del complejo galáctico y sus impactos en el nivel de la ETI con valores variables de parámetros clave. Nuestro modelo cubre toda la galaxia, lo que nos permite descartar la restricción de su rango limitado de radio galáctico del trabajo anterior [ 19]; una restricción impidió que su modelo obtuviera una ubicación de pico precisa, y su resultado estaba más allá del límite inferior del rango (<2.5 kpc).
Observamos que es difícil imponer una restricción adicional al número de ETI debido a la incertidumbre inherente de cualquier valor de parámetro específico; sin embargo, nuestras principales conclusiones no se modifican. Cuando se apliquen más análisis futuros sobre la aparición y el desarrollo de ETI, podemos desarrollar un rango mejorado para la cantidad de ETI con varianza espacial y temporal. También notamos que nuestra simulación de gas y conversión de gas a estrellas es burda, con una tabla hash espacial estática de la Vía Láctea que carece de las consideraciones de la cinemática estelar; sin embargo, solo nos enfocamos en la propensión al crecimiento de ETI en una escala galáctica, que no está fuertemente asociada a la migración de estrellas.
4. Conclusiones
Se ha desarrollado un modelo galáctico con resolución de planetas individuales similares a la Tierra para analizar dónde y cuándo la vida inteligente potencial es más abundante a lo largo del tiempo, y qué edad tiene la mayoría de la vida compleja en el momento actual en la Vía Láctea. Se forma un análisis preciso para la propensión de ETI potencial, con un conjunto de criterios que incluyen: la formación de dos fases distintas de la Galaxia, análisis observacional reciente sobre la prevalencia de planetas similares a la Tierra, esterilización de SNe que amenazan la vida terrestre, Proceso de Poisson de abiogénesis, diferentes escalas de tiempo evolutivas y posible autoaniquilación de la vida. Utilizamos tres parámetros, λ A , T evo , P ann, rastrear el origen, evolución y desarrollo de la vida en la Vía Láctea, y evaluar las variaciones espaciales y temporales de la cantidad de ETI.
La extensión del tiempo a 20 Gyrs, independientemente del cambio en los parámetros clave, dio como resultado un pico para el número de ETI en todas las regiones galácticas alrededor de 4 kpc, y el tiempo en 8 Gyrs. El número de ETI disminuyó espacial y temporalmente desde el pico. Incluso si variamos la escala de tiempo evolutiva para que sea 2 Gyrs menor o mayor de lo que se requería en la Tierra, y con diferentes valores de λ A y escenarios de casos extremos de P ann(0 y 0,99), nuestra conclusión de la ubicación precisa y la hora del pico no se modifica. Nuestros resultados sugieren que la cantidad de vida inteligente no siempre aumenta con el tiempo; de hecho, nuestro modelo predice que después de que ocurre el pico, el número de ETI comienza a disminuir monótonamente con el tiempo, y esta propensión permanece durante los siguientes 6.5 Gyrs. Además, nuestros resultados muestran que el nivel de ETI eventualmente alcanzará un equilibrio entre el nacimiento y la muerte de la vida inteligente en aproximadamente 20 Gyrs. Una investigación adicional sobre el número de eventos de esterilización de SNe sugiere que la prevalencia de ETI alcanza su máximo a pesar de tener la frecuencia más alta de eventos de esterilización de SNe por la ubicación del pico.
También llegamos a la conclusión de que en el momento actual del estudio, la mayoría de la vida inteligente en la Galaxia es menor de 0,5 Gyr, con valores del parámetro de probabilidad de autoaniquilación entre 0 y 0,01; con un valor relativamente más alto del parámetro de aniquilación (0.1), la vida más inteligente es menor de 0.01 Gyr. Como no podemos asumir una baja probabilidad de aniquilación, es posible que la vida inteligente en otras partes de la Galaxia sea todavía demasiado joven para ser observada por nosotros. Por lo tanto, nuestros hallazgos pueden implicar que la vida inteligente puede ser común en la Galaxia pero aún es joven, lo que respalda el aspecto optimista para la práctica de SETI. Nuestros resultados también sugieren que nuestra ubicación en la Tierra no está dentro de la región donde se asienta la mayor parte de la vida inteligente, y las prácticas SETI deben estar más cerca del interior de la Galaxia, preferiblemente en el anillo a 4 kpc del Centro Galáctico.
Además, encontramos que la autoaniquilación potencial es muy influyente en la cantidad de vida inteligente galáctica, lo que sugiere otra posible respuesta a la paradoja de Fermi; si es probable que la vida inteligente se destruya a sí misma, no es de extrañar que haya poca o ninguna vida inteligente en otros lugares. Además, no se encontró que la probabilidad de abiogénesis influyera en la cantidad de vida inteligente dentro del rango desarrollado por trabajos anteriores, lo que apoya la perspectiva optimista de que la vida es común en la Galaxia.
Los resultados de nuestro modelo arrojan una gama de posibles cantidades para la vida inteligente a lo largo del tiempo. Afirmamos que la vida inteligente siempre será más abundante a aproximadamente 4 kpc desde el centro galáctico, alcanzando su punto máximo alrededor de los 8 Gyrs, disminuyendo monótonamente desde ese punto máximo, y que la mayoría de la vida inteligente potencial aún es joven. El número exacto de vida inteligente estimado aquí no es el foco de nuestro trabajo; más bien, es el desarrollo de una imagen galáctica completa y estadística que rastrea la propensión potencial al crecimiento de la vida inteligente en un curso de ~ 20 mil millones de años.
Contribuciones de autor
Conceptualización, JHJ; metodología, XC y JHJ; software, XC y JHJ; validación, JHJ, KAF y YLY; análisis formal, XC y JHJ; investigación, XC y JHJ; recursos, JHJ; curación de datos, XC y JHJ; redacción: preparación del borrador original, XC y JHJ; redacción: revisión y edición, JHJ, KAF y YLY; visualización, XC y JHJ; supervisión, JHJ; administración de proyectos, JHJ; Adquisición de fondos, JHJ y YLY Todos los autores han leído y aceptado la versión publicada del manuscrito.
Fondos
Esta investigación fue financiada por el Programa de Investigación de Exoplanetas de la NASA NNH18ZDA001N-2XRP.
Declaración de la Junta de Revisión Institucional
Este artículo ha sido aprobado para su publicación ilimitada por los Servicios de Revisión de Documentos del Laboratorio de Propulsión a Chorro (JPL), Instituto de Tecnología de California. Registro: URS297443; Enviado por el autor: Jonathan H Jiang el 13/12/2020; Tipo de documento: Artículo de revista; Título: Una estimación estadística de la presencia de inteligencia extraterrestre en la Vía Láctea; El número de autorización JPL es CL # 21-0186.
Declaración de consentimiento informado
No aplica.
Declaración de disponibilidad de datos
Los datos subyacentes a este artículo están disponibles en el artículo. Si tiene preguntas adicionales sobre el intercambio de datos, comuníquese con el autor correspondiente en Jonathan.H.Jiang@jpl.nasa.gov .
Expresiones de gratitud
Esta investigación fue apoyada por el Laboratorio de Propulsión a Chorro del Instituto de Tecnología de California, en virtud del contrato con la NASA. Agradecemos el apoyo financiero parcial del Programa de Investigación de Exoplanetas de la NASA NNH18ZDA001N-2XRP. También agradecemos el apoyo del Laboratorio Planetario Virtual de la Universidad de Washington.
Conflictos de interés
Los autores declaran no tener ningún conflicto de intereses.
Referencias
- Tarter, J. La búsqueda de inteligencia extraterrestre (SETI). Annu. Rev. Astron. Astrophys. 2001 , 39 , 511–548. [ Google Académico ] [ CrossRef ]
- Hart, MH Explicación de la ausencia de extraterrestres en la Tierra. QJR Astron. Soc. 1975 , 16 , 128. [ Google Scholar ]
- Carroll-Nellenback, J .; Frank, A .; Wright, J .; Scharf, C. La paradoja de Fermi y el efecto Aurora: asentamiento, expansión y estados estacionarios de exo-civilización. Astron. J. 2019 , 158 , 117. [ Google Académico ] [ CrossRef ]
- Foucher, F .; Hickman-Lewis, K .; Westall, F .; Brack, A. Un enfoque estadístico para ilustrar el desafío de la astrobiología para la divulgación pública. Life 2017 , 7 , 40. [ Google Académico ] [ CrossRef ] [ PubMed ]
- Drake, FD La búsqueda por radio de vida extraterrestre inteligente. En Aspectos actuales de la exobiología ; Mamikunian, G., Briggs, MH, Eds .; Oxford University Press: Oxford, Reino Unido, 1965; págs. 323–345. [ Google Académico ]
- Walters, C .; Hoover, RA; Kotra, RK Colonización interestelar: ¿Un nuevo parámetro para la ecuación de Drake? Ícaro 1980 , 41 , 193-197. [ Google Académico ] [ CrossRef ]
- Burchell, MJ ¿W (h) ither la ecuación de Drake? En t. J. Astrobiol. 2006 , 5 , 243–250. [ Google Académico ] [ CrossRef ]
- Forgan, DH Un banco de pruebas numérico para hipótesis de vida e inteligencia extraterrestres. En t. J. Astrobiol. 2009 , 8 , 121-131. [ Google Académico ] [ CrossRef ]
- Cirkovic, MM Earths: ¿Raras en el tiempo, no en el espacio? J. Br. Interplaneta. Soc. 2004 , 57 , 53. [ Google Scholar ]
- Maccone, C. La ecuación estadística de Drake. Acta Astronaut. 2010 , 67 , 1366-1383. [ Google Académico ] [ CrossRef ]
- Glade, N .; Ballet, P .; Bastien, O. Un enfoque de proceso estocástico de los parámetros de la ecuación de Drake. En t. J. Astrobiol. 2012 , 11 , 103–108. [ Google Académico ] [ CrossRef ]
- Balazs, B. El cinturón galáctico de la vida inteligente. En bioastronomía: los próximos pasos ; Springer: Dordrecht, Holanda, 1988; págs. 61–66. [ Google Académico ]
- González, G .; Brownlee, D .; Ward, P. La zona habitable galáctica: evolución química galáctica. Ícaro 2001 , 152 , 185–200. [ Google Académico ] [ CrossRef ]
- Lineweaver, CH; Fenner, Y .; Gibson, BK La zona habitable galáctica y la distribución por edades de la vida compleja en la Vía Láctea. Science 2004 , 303 , 59–62. [ Google Académico ] [ CrossRef ] [ PubMed ]
- Gowanlock, MG; Patton, DR; McConnell, S. Un modelo de habitabilidad dentro de la Vía Láctea. Astrobiology 2011 , 11 , 855–873. [ Google Académico ] [ CrossRef ] [ PubMed ]
- Naab, T .; Ostriker, JP Un modelo simple para la evolución de las galaxias de disco: La Vía Láctea. Lun. No. R. Astron. Soc. 2006 , 366 , 899–917. [ Google Académico ] [ CrossRef ]
- Jurić, M .; Ivezić, Ž .; Brooks, A .; Lupton, RH; Schlegel, D .; Finkbeiner, D .; Padmanabhan, N .; Bond, N .; Sesar, B .; Rockosi, CM; et al. Tomografía de la Vía Láctea con SDSS. I. distribución de densidad de números estelares. Astrophys. J. 2008 , 673 , 864–914. [ Google Académico ] [ CrossRef ]
- Prantzos, N. Sobre la “zona habitable galáctica”. Ciencia espacial. Rev. 2008 , 135 , 313–322. [ Google Académico ] [ CrossRef ]
- Morrison, IS; Gowanlock, MG Ampliación del modelado de zonas habitables galácticas para incluir el surgimiento de vida inteligente. Astrobiology 2015 , 15 , 683–696. [ Google Académico ] [ CrossRef ]
- Forgan, DH; Rice, K. Prueba numérica de la hipótesis de las tierras raras utilizando técnicas de realización de Monte Carlo. En t. J. Astrobiol. 2010 , 9 , 73–80. [ Google Académico ] [ CrossRef ]
- Hair, TW Dispersión temporal de la emergencia de la inteligencia: un análisis del tiempo entre llegadas. En t. J. Astrobiol. 2011 , 10 , 131-135. [ Google Académico ] [ CrossRef ]
- Spiegel, D .; Turner, E. Análisis bayesiano de las implicaciones astrobiológicas de la aparición temprana de la vida en la Tierra. Proc. Natl. Acad. Sci. Estados Unidos 2012 , 109 , 395–400. [ Google Académico ] [ CrossRef ]
- Scharf, C .; Cronin, L. Cuantificando los orígenes de la vida a escala planetaria. Proc. Natl. Acad. Sci. EE. UU. 2016 , 113 , 8127–8132. [ Google Académico ] [ CrossRef ] [ PubMed ]
- Chen, J .; Kipping, D. Sobre la tasa de abiogénesis desde una perspectiva informática bayesiana. Astrobiología 2018 , 18 , 1574-1584. [ Google Académico ] [ CrossRef ] [ PubMed ]
- Kipping, D. Un análisis bayesiano objetivo del comienzo temprano de la vida y nuestra llegada tardía. Proc. Natl. Acad. Sci. USA 2020 , 117 , 11995–12003. [ Google Académico ] [ CrossRef ] [ PubMed ]
- Livio, M. ¿Qué tan raras son las inteligencias extraterrestres y cuándo surgieron? Astrophys. J. 1999 , 511 , 429–431. [ Google Académico ] [ CrossRef ]
- Carter, B. ¿Escenario de evolución de cinco o seis pasos? En t. J. Astrobiol. 2008 , 7 , 177–182. [ Google Académico ] [ CrossRef ]
- Salpeter, EE La función de luminosidad y evolución estelar. Astrophys. J. 1955 , 121 , 161-167. [ Google Académico ] [ CrossRef ]
- Petigura, EA; Howard, AW; Marcy, GW Prevalencia de planetas del tamaño de la Tierra que orbitan estrellas similares al Sol. Proc. Natl. Acad. Sci. EE. UU. 2013 , 110 , 19273–19278. [ Google Académico ] [ CrossRef ]
- Hastings, EJ; Mesit, J .; Guha, RK Optimización de simulaciones en tiempo real a gran escala mediante hash espacial. En Actas de la Conferencia de verano de simulación por computadora de 2005, Cherry Hill, Nueva Jersey, EE. UU., 24-28 de julio de 2005; Parte de la multiconferencia de simulación de verano de 2005 (SummerSim'05). Curran Associates, Inc .: Red Hook, Nueva York, EE. UU., 2005; págs. 9-17. [ Google Académico ]
- Fuentes, SA; De Ridder, J .; Debosscher, J. Identificación de la estructura de densidad jerárquica del halo estelar usando (F) OPTICS. Astron. Astrophys. 2017 , 599 , A143. [ Google Académico ] [ CrossRef ]
- Donato, F .; Gentile, G .; Salucci, P .; Martins, CF; Wilkinson, MI; Gilmore, G .; Grebel, EK; Koch, A .; Wyse, R. Una densidad constante de la superficie del halo de materia oscura en las galaxias. Lun. No. R. Astron. Soc. 2009 , 397 , 1169-1176. [ Google Académico ] [ CrossRef ]
- Schmidt, M. La tasa de formación de estrellas. Astrofísica 1959 , 129 , 243. [ Google Scholar ] [ CrossRef ]
- Kennicutt, RC, Jr. La ley global de Schmidt en las galaxias formadoras de estrellas. Astrophysics 1998 , 498 , 541–552. [ Google Académico ] [ CrossRef ]
- Arifyanto, MI; Fuchs, B .; Jahreiß, H .; Wielen, R. Cinemática de estrellas subenanas cercanas. Astron. Astrophys. 2005 , 433 , 911–916. [ Google Académico ] [ CrossRef ]
- Hansen, CJ; Kawaler, SD Stellar Interiors: Principios físicos, estructura y evolución ; Springer: Belin, Alemania, 1994. [ Google Scholar ]
- Sackmann, IJ; Boothroyd, AI; Kraemer, KE Our Sun. III. Presente y futuro. Astrophys. J. 1993 , 418 , 457–468. [ Google Académico ] [ CrossRef ]
- Snaith, O .; Haywood, M .; Matteo, P .; Lehnert, M .; Combes, F .; Katz, D .; Gómez, A. Reconstrucción de la historia de formación estelar de los discos de la Vía Láctea a partir de abundancias químicas. Astron. Astrophys. 2014 , 578 , A18. [ Google Académico ] [ CrossRef ]
- Gilmore, G .; Wyse, RFG La evolución química de la galaxia. Nature 1986 , 322 , 806–807. [ Google Académico ] [ CrossRef ]
- Hopkins, AM; Beacom, JF Sobre la normalización de la historia de la formación de estrellas cósmicas. Astrophys. J. 2006 , 651 , 142-154. [ Google Académico ] [ CrossRef ]
- Madau, P .; Dickinson, M. Historia de la formación de estrellas cósmicas. Annu. Rev. Astron. Astrophys. 2014 , 52 , 415–486. [ Google Académico ] [ CrossRef ]
- Fenner, Y .; Gibson, BK Derivación de la función de distribución de la metalicidad de los sistemas galácticos. PASA 2003 , 20 , 189-195. [ Google Académico ] [ CrossRef ]
- Tammann, G .; Loeffler, W .; Schroeder, A. La tasa de supernova galáctica. Astrophys. J. Supl. Ser. 1994 , 92 , 487–493. [ Google Académico ] [ CrossRef ]
- Tutukov, AV; Yungelson, LR; Iben, I., Jr. Las frecuencias de las supernovas en binarios. Astrophys. J. 1992 , 386 , 197-205. [ Google Académico ] [ CrossRef ]
- Meng, X .; Yang, W. Un modelo progenitor integral para SNe Ia. Astrophys. J. 2010 , 710 , 1310-1323. [ Google Académico ] [ CrossRef ]
- Gehrels, N .; Laird, CM; Jackman, CH; Cannizzo, JK; Mattson, BJ; Chen, W. Agotamiento del ozono por supernovas cercanas. Astrophys. J. 2003 , 585 , 1169-1176. [ Google Académico ] [ CrossRef ]
- Miller, D .; Branch, D. Distribuciones de magnitud absoluta de supernova. Astrophys. J. 1990 , 100 , 530. [ Google Scholar ] [ CrossRef ]
- Richardson, D .; Jenkins, R .; Wright, J .; Maddox, L. Distribuciones de magnitud absoluta de supernovas. Astron. J. 2014 , 147 , 118. [ Google Scholar ] [ CrossRef ]
- Wesson, P. Panspermia, pasado y presente: condiciones astrofísicas y biofísicas para la diseminación de la vida en el espacio. Ciencia espacial. Rev. 2010 , 156 , 239–252. [ Google Académico ] [ CrossRef ]
- Hoerner, SV La búsqueda de señales de otras civilizaciones. Science 1961 , 134 , 1839–1843. [ Google Académico ] [ CrossRef ] [ PubMed ]
- Nick, B. Riesgos existenciales analizando escenarios de extinción humana y peligros relacionados. J. Evol. Technol. 2002 , 9 . [ Google Académico ]
- Billings, L. Alien Anthropocene: ¿Cómo lucharían otros mundos contra el cambio climático? Sci. Soy. 2018 , 28 . [ Google Académico ]
- Sotos, JG Biotechnology y la vida de las civilizaciones técnicas. En t. J. Astrobiol. 2019 , 18 , 445–454. [ Google Académico ] [ CrossRef ]
- Dutil, Y .; Dumas, S. Sostenibilidad: un camino tedioso hacia la colonización galáctica. arXiv 2007 , arXiv: 0711.1777. [ Google Académico ]
- Haqq-Misra, JD; Baum, SD La solución sostenible a la paradoja de Fermi. arXiv 2009 , arXiv: 0906.0568. [ Google Académico ]
- Grinspoon, D. Planetas solitarios, La filosofía natural de la vida extraterrestre ; HarperCollins Publishers Inc .: Nueva York, NY, EE. UU., 2003. [ Google Scholar ]
- Nittler, LR Buscando vida en lugares lejanos. Science 2004 , 303 , 1614. [ Google Scholar ] [ CrossRef ]
- Forgan, DH; Dayal, P .; Cockell, C .; Noam, L. Evaluación de la habitabilidad galáctica utilizando simulaciones cosmológicas de alta resolución de la formación de galaxias. arXiv 2015 , arXiv: 1511.01786. [ Google Académico ] [ CrossRef ]
- Ryder, G. Bombardeo de la Tierra Hadeana: ¿Saludable o perjudicial? Astrobiology 2003 , 3 , 3. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Schmieder, M .; Krin, DA Eventos de impacto de la Tierra a través del tiempo geológico: una lista de edades recomendadas para estructuras y depósitos de impacto terrestre. Astrobiology 2020 , 20 , 91. [ Google Scholar ] [ CrossRef ]
Nota del editor: MDPI se mantiene neutral con respecto a los reclamos jurisdiccionales en mapas publicados y afiliaciones institucionales. |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
No hay comentarios:
Publicar un comentario